Visione ed esplorazione visuo-spaziale
Gran parte (oltre il 60%) dell’attività cerebrale è connessa con l’elaborazione del segnale visivo.
Estese aree corticali, dalle regioni occipitali a quelle frontali e sottocorticali, sono implicate nei processi
visivi che vanno dalla percezione del segnale sensoriale alla sua successiva elaborazione ed integrazione per
la programmazione di risposte motorie o per l’acquisizione di complesse esperienze cognitive.
Circa il 90% del tempo impiegato nella visione è dedicato alla fissazione il rimanente 10% è usato per spostare gli
occhi dall’attuale punto di fissazione ad un punto successivo. In media avvengono circa tre fissazioni al secondo durante
la visione attiva in questi casi ogni fissazione ha una durata variabile (300-500 ms).
Le fissazioni sono intercalate da movimenti rapidi degli occhi che consentono lo spostamento dello sguardo da un punto di interesse al successivo.
Questi rapidi movimenti di “gaze shift” sono chiamati saccadi ed hanno una velocità che può superare i 700°/s . La percezione e l’elaborazione del
segnale visivo è dipendente ed a sua volta modula il programma motorio necessario allo spostamento degli occhi per l’esplorazione visiva.
L’architettura anatomo-funzionale del sistema visivo è strettamente interconnessa ed è mantenuta dalla retina alle aree corticali con estrema precisione.
Tale architettura consente al cervello di ottenere la maggiore fonte di informazioni necessarie per l’orientamento spaziale.
La risoluzione spaziale ottimale di un oggetto o di un dettaglio avviene in un’area specifica della retina chiamata fovea. La fovea ha un diametro di 1.5 mm
contiene la maggiore densità di fotorecettori (trasduttori del segnale luminoso in segnale elettrico) con capacità di discriminare minimi dettagli spaziali
e specifiche lunghezze d’onda spettrali relative ai principali colori.
La risoluzione spaziale decresce drammaticamente procedendo dalla fovea verso la periferia retinica dove invece aumenta progressivamente la risoluzione temporale
ovvero la capacità di discriminare il movimento. Questa caratteristica funzionale dipende non soltanto dalla densità dei fotorecettori per superficie retinica,
ma anche dal tipo prevalente di fotorecettore presente nel centro e nella periferia. I coni sono distribuiti prevalentemente nella regione foveale mentre i bastoncelli nella periferia.
Proprio a causa delle sue caratteristiche anatomo funzionali, il sistema visivo presenta una fisiologica limitata capacità di processazione degli stimoli
sensoriali rispetto alla quantità di dettagli che provengono dall’ambiente esterno. Ciò comporta la necessità di ridurre ovvero: “filtrare” l’eccesso
d’informazioni ambientali, selezionando gli stimoli percettivi più salienti.
Secondo quali criteri avviene questa selezione? Varie teorie sono state proposte negli ultimi anni per spiegare quali siano i meccanismi che guidano
la selezione durante l’esplorazione visuo-spaziale. Alcune teorie preattentive enfatizzano gli effetti delle caratteristiche fisiche dell’immagine
(colore, forma, contrasto ed orientamento) sulla percezione visiva e quindi sulla direzione dello sguardo durante l’esplorazione visuospaziale.
In questo caso lo spostamento degli occhi sarebbe guidato dalla salienza dell’immagine secondo un meccanismo bottom-up.
Altre ipotesi sottolineano l’importanza del controllo cognitivo (goal-driven) sulla direzione di sguardo in relazione alle richieste del task
seguendo un meccanismo top-down. I dati più attuali tendono ad unificare entrambe le ipotesi suggerendo che inizialmente il sistema visivo
utilizza un livello di selezione “bottom up” dell’immagine basato sulle proprietà fisiche della stessa (orientamento, colore, contrasto).
Queste caratteristiche bottom-up possono essere riprodotte attraverso una mappa di salienza che corrisponde alla rappresentazione percettiva di
ciascuna immagine nella corteccia visiva. Le parti più salienti dell’immagine competono tra loro per catturare la fissazione successiva secondo
un modello di esplorazione visiva così detto “winner take all”. Questo sistema sembra essere preponderante durante le fasi iniziali dell’ esplorazione
visuospaziale, successivamente, man mano che l’elaborazione cognitiva aumenta, l’esplorazione visiva è guidata da meccanismi top-down (goal driven).
A questo punto è abbastanza chiaro che la selezione e l’elaborazione dello stimolo visivo è strettamente connessa alla programmazione del movimento
oculare successivo. In altre parole prima ancora che gli occhi incomincino a muoversi, l’attenzione percettiva si sposta sul bersaglio della saccade
futura. Secondo questa ipotesi nota come “teoria premotoria” dell’attenzione, esisterebbe una stretta correlazione tra meccanismi che regolano la
selezione attentiva e quelli che determinano il programma oculo-motorio. Numerosi studi di neurofisiologia, neuro immagini e registrazione di attività
neuronale nelle scimmie, hanno maggiormente chiarito l’ipotesi premotoria, confermando l’esistenza di una sovrapposizione anatomo funzionale tra importanti
networks occipito-parieto-frontali, implicati nella percezione visiva , nell’orientamento attentivo e nella programmazione dei movimenti oculari.
Una serie di eleganti e rilevanti studi di Sommer & Wurz, hanno inoltre definitivamente chiarito l’esistenza di un via che integra la direzione
dell’attenzione con la programmazione del movimento oculare. In particolare essi hanno dimostrato l’esistenza di una via collicolo talamica
(nucleo dorso-mediale del talamo) che è importante nel remapping spaziale degli oggetti di una scena, durante i movimenti saccadici.
Questa via raggiunge neuroni visuali delle aree frontali (FEF) e probabilmente i neuroni parietali posteriori (LIP), determinando in anticipo
uno un shift nella risposta dall’attuale campo recettivo neuronale, al campo recettivo futuro, quello cioè che occuperebbe alla fine della
saccade successiva. Queste importanti scoperte hanno permesso non soltanto di comprendere meglio come lo sguardo si orienta nello spazio,
ma anche come la visione del mondo circostante sia mantenuta stabile durante i movimenti degli occhi.

Figura 1 - Visione foveale. Nella fovea avviene la maggiore discriminazione spaziale degli oggetti, massima acutezza visiva.
Un'altra importante caratteristica dell’esplorazione visiva è che durante la scannerizzazione di una scena di interesse, gli occhi si muovono
attraverso una sequenza di brevi fissazioni all’interno di piccole regioni d’interesse, durante questa processo, lo sguardo non è mai a riposo, anzi,
gli occhi sono in continuo micromovimento. Questi micromovimenti inconsapevoli, chiamati “microsaccadi” sono necessari per la percezione e dipendono
dalle caratteristiche fisiologiche dei neuroni della corteccia visiva primaria (V1), in assenza di micromovimenti l’immagine diventerebbe immediatamente
sfuocata dopo la sua percezione.
E’ noto che i micromovimenti oculari associati alla fissazione, detti microsaccadi, aumentano durante lo scanning di aree a maggiore salienza informativa.
Ciò consente di ritenere un maggior numero di dettagli dell’oggetto esaminato. In altre parole, maggiore è il tempo impiegato nella fissazione, maggiore
è il grado di attenzione e più elaborati sono i processi cognitivi associati. Le microsaccadi, benché siano dei movimenti miniaturizzati difficilmente
differenziabili dal rumore, con un’ampiezza di circa 1° e frequenza di 1 movimento al secondo durante la fissazione, hanno le stesse caratteristiche
dinamiche delle saccadi maggiori. Le microsaccadi sembrerebbero indicare l’orientamento verso cui è diretta l’attenzione (shift dell’attenzione) e
possono essere usate per studiare come, il sistema oculomotorio, si prepara a dirigere la fissazione verso l’oggetto di interesse.
Classificazione dei movimenti oculari
Diverse classi di movimenti oculari sono necessarie non soltanto per stabilizzare l’immagine sulla retina durante le rotazioni del capo o durante gli
spostamenti dell’immagine stessa in movimento, ma anche per spostare lo sguardo repentinamente su un nuovo punto d’interesse. Da un punto di vista
funzionale si può dire che nell’uomo sono presenti due principali tipi di movimenti oculari: quelli che stabilizzano lo sguardo (nistagmo optocinetico,
riflesso oculovestibolare) e quelli che spostano lo sguardo (saccadi, pursuit, vergenze).
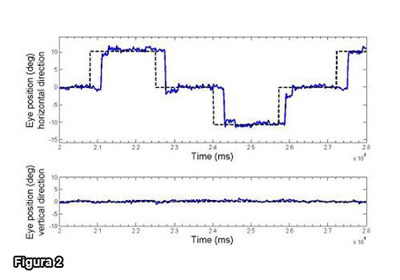
Figura 2 - Saccadi orizzontali con ampiezza di 10°.
Molti dei substrati anatomo-funzionali alla base di queste sottoclassi di movimenti oculari sono noti, anche se senza dubbio i più studiati sono i
movimenti saccadici. In condizioni naturalistiche, tuttavia i vari tipi di movimenti oculari si succedono in modo automatico e consentono uno
“scanning” continuo della scena visiva. La sequenza di fissazioni/ non fissazioni, durante l’esplorazione visiva è espressione di numerosi processi
cognitivi; l’utilizzo di task controllati e ripetibili, in cui è possibile manipolare le variabili (spazio/temporali) consente di esaminare specifici
domini cognitivi (es percezione, attenzione, memoria, motivazione, finalizzazione). Da un punto di vista generale, i movimenti oculari sono in larga parte
rappresentati da circuiti con feedback in cui il comando motorio può essere grossolanamente riassunto in tre tipologie: Volontario-Involontario-Riflesso.
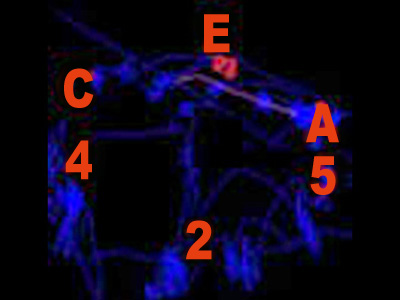
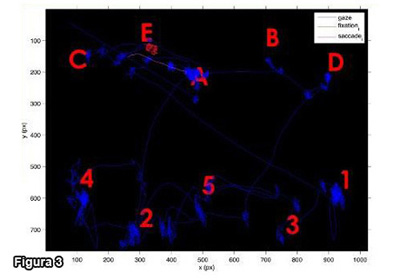 Figura 3 - Esplorazione di immagine cognitivamente rilevante (Veneri & Rufa 2011).
Figura 3 - Esplorazione di immagine cognitivamente rilevante (Veneri & Rufa 2011).
Questi segnali sono integrati a livello sottocorticale da un integratore neurale per i movimenti orizzontali ed uno separato per i movimenti verticali.
La funzione dell’integratore è di trasformare il segnale di velocità in un segnale di posizione degli occhi. A questo livello, in considerazione delle
diverse sottoclassi di movimenti oculari, numerose strutture del tronco encefalico, del cervelletto e dei gangli della base partecipano all’integrazione.
L’output del sistema è condotto ai muscoli extraoculari per il raggiungimento ed il mantenimento della posizione desiderata.
 Figura 4 - Esplorazione visiva con una tecnica di gaze contingent che consente di vedere nitidamente soltanto nella parte centrale (visione foveale)
mentre la periferia è parzialmente o totalmente oscurata (Veneri & Rufa 2011).
Figura 4 - Esplorazione visiva con una tecnica di gaze contingent che consente di vedere nitidamente soltanto nella parte centrale (visione foveale)
mentre la periferia è parzialmente o totalmente oscurata (Veneri & Rufa 2011).
Conclusioni
In conclusione la comprensione dei processi visivi e della loro elaborazione, lo studio della percezione visiva e dell’attenzione visuo-spaziale e
l’identificazione dei processi che sottendono l’elaborazione e l’esecuzione del programma oculomotorio hanno da sempre costituito un obiettivo primario
nelle neuroscienze di base e cognitive. Essi inoltre hanno rappresentato da sempre una importante sfida per gli studiosi dei meccanismi cerebrali.
I molti progressi che sono stati fatti negli ultimi anni in questo settore non soltanto hanno consentito di definire meglio il funzionamento dei network
visivi e visuomotori, ma più in generale hanno consentito di aggiungere alcuni fondamentali tasselli per comprendere i meccanismi generali del funzionamento
del nostro cervello.
Prof.ssa Alessandra Rufa, MD, PhD
Eye-tracking & Visual Applications Lab (EVALab)
Dipartimento di Scienze Neurologiche Neurochirurgiche e del Comportamento
Università degli Studi di Siena
www.evalab.unisi.it
Letture consigliate
- Awh, E., Armstrong, K. M., & Moore, T. Visual and oculomotor selection: links, causes and implications for spatial attention. Trends Cogn Sci, 2006;10(3): 124–130.
- Büttner-Ennever JA. Mapping the oculomotor system. Prog Brain Res 2008;171:3-11.
- Findlay, J. M., Brown, V., & Gilchrist, I. D.. Saccade target selection in visual search: The effect of information from the previous fixation. Vision Res. 2001; 41(1): 87–95.
- Gilchrist, I. D., Heywood, C. A., & Findlay, J. M. (, Apr). Saccade selection in visual search. Vision Res. 1999; 39(7):1373–1383.
- Leigh R.J., Zee. D. The neurology of eye movements (book/dvd). fourth edition. 2006 New York: Oxford University Press.
- Mariottini GL, Prattichizzo D, De Biasi M, Snickars C, Rufa A, De Capua A, Rossi S. Human robotic interface for the interaction in cognitive and emotional huma domains.IEEE/RSJ. International Conference on Intelligent Robots and Systems. S Diego,CA,USA, Oct 29-nov 2. 2007
- Nobre AC, Sebestyen GN, Gitelman DR,Mesulam MM, Frackowiak RS, Frith CD. Functional localization of the system for visuospatial attention using positron emission tomography. Brain 1997;120:515-533
- Pomplun, M., Reingold, E. M., & Shen, J. Peripheral and parafoveal cueing and masking effects on saccadic selectivity in a gaze-contingent window paradigm. Vision Research. 2001
- Rizzolatti G, Craighero L. Spatial attention: mechanisms and theories. In Advances in Psychological Sciences Vol.2: Biological and Cognitive Aspects (M Sabourin, F Craik et al. Eds). Pp171-198. Psychology Press, Hove, England
- Rayner, K. Eye movements in reading and information processing: 20 years of research. Psychol Bull.1998;124(3), 372–422.
- Robinson DA, Oculomotor control signals. In: Basic mechanisms of ocular motility and their clinical implications. Lennerstrand G, Bach-y-Rita P, , ed., 1975, pp 337–374. Pergamon Press, Oxford.
- Robinson FR, Fuchs AF, Noto CT. Cerebellar influences on saccade plasticity. Ann N Y Acad Sci. 2002;956:155-63.
- Rufa A, Mariottini GL, Prattichizzo D, Alessandrini D, Vicino A, Federico A. Videobased eye-tracking: our experience with advanced stimuli design foe eye tracking software. Ann NY Acad Sci 2005;1039:575-9
- Scudder CA, Moschovakis AK, Karabelas AB, Highstein SM, Anatomy and physiology of saccadic long-lead burst neurons recorded in the alert squirrel monkey. II. Pontine neurons, Journal of Neurophysiology. 1996. 76(1):353-370.
- Sommer MA, Wurtz RH. Visual perception and corollary discharge. Perception 2008; 37(3):408-18
- Yantis S. the control of visual attention. In H Pashler (Ed) 1998. Attention, pp 223-256). East Sussex, UK:Psychology Press.
- Veneri G, Federighi P, Rosini F, Federico A and Rufa A. Influences of data filtering on human computer interaction by gaze-contingent display and eye-tracking applications. Comp.Hum. Behav.2011
- Veneri G, Federighi P, Pretegiani E, Rosini F, Federico A and Rufa A. EVA: Eye Tracking Stimulus Integrated Semi Automated Case Base System. The 13th World Multi-Conference on Systemics, Cybernetics and Informatics.WMSCI July 10-13,2009 Orlando, Florida USA.
- Yarbus, A. L. Eye movements and vision. (Vol. chapter VII; e. L. A. Riggs, Ed.). 1967.Plenum Press, New York.
- Wright RD&Ward LM. Orienting of attention. 2008, Oxford University Press
- Wurtz RH. Neuronal mechanism of visual stability. Vis Res 2008;48,20:2070-89.